![]()
|
Man has a yetzer hara (evil inclination) and a yetzer tov (good inclination). The existence of the two allows man to have the free will to choose good. The yetzer hara will first try to grab a person through a subtle gesture. If an individual falls prey at the early stage he will likely fall deeper and deeper into it's clutches. This process is similar to one which exits in nature (see article below). There is a wasp that stings a spider, lays its eggs on it, gets it to spin a totally different web, the eggs then hatch and eat the spider alive. The goal is to overcome the yetzer hara on its onset because this is when we are strongest and most able to resist its pull.
NATURE | VOL 406 | 20 JULY 2000 |
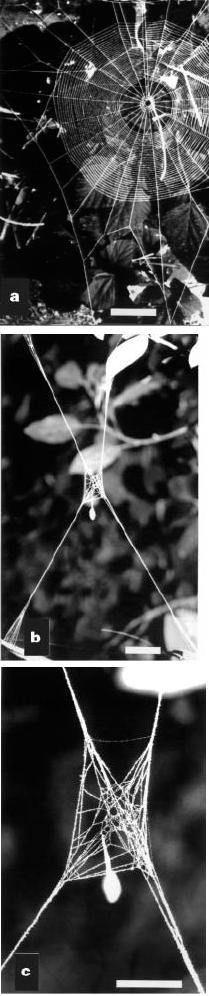
www.nature.com A parasitic wasp forces its host to weave The female Hymenoepimecis sp. wasp attacks the spider Plesiometa argyra at the hub of its orb, stings it into temporary paralysis and lays an egg on the spider�s abdomen [1]. Subsequently, the spider resumes normal activity. During the next 7�14 days, it builds apparently normal orbs (Fig. 1a) to capture prey, while the wasp�s egg hatches and the larva grows by sucking the spider�s haemolymph. On the night that it will kill its host, the larva induces the spider to build a cocoon web, moults, kills and consumes the spider, and then spins its pupal cocoon hanging by a line from the cocoon web. Apparently undamaged cocoon webs almost always had several lines (mean, 3.851.4; range, 2�8; n439) radiating in a plane from a �hub� (Fig. 1b�f). Most radial lines branched near their tips and were attached to the substrate at many points (Fig. 1b). Most webs had only radii and lines connecting them at the hub, and lacked circular hub lines (86% of 66) or frame lines (65% of 77) connecting the radial lines. Any frame lines present were typically much shorter and nearer the hub than those of normal orbs (Fig. 1e). The central portion of the hub was never empty, as it is in a normal orb (Fig. 1a). The most elaborate cocoon webs, however, had distinct hub loops, frames, and a mesh above and below the hub (Fig. 1g), confirming that they were modified orbs. The construction of cocoon webs was very consistent, and it appeared to be the same as the early stages of type �D� frame construction (ref. 2, and W.G.E., manuscript in preparation). Many other integral features of normal orb-web construction, including breaking and then reeling up and replacing lines, and breaking and then reattaching lines [2�6], were completely absent. A single �mistake� in this respect by the spider could be disastrous for the wasp larva, as the many-stranded cable of radial lines would be replaced with a single pair of drag lines. The temporary and sticky spirals were also missing, and the central portion of the finished hub was not removed [2�6]. These differences between cocoon webs and normal orbs all make the cocoon web a stronger, more durable support for the wasp�s cocoon. The importance of this is illustrated by the vulnerability of pupae of the related wasp H. robertsae to heavy rains [7]. The larva makes small holes in the spider�s abdomen to imbibe haemolymph1. As the spider continues to build the cocoon web even when the larva is removed shortly before construction would normally start, the changes in the spider�s behaviour must be induced chemically rather than by direct physical interference. The effects are both rapid (removal earlier in the evening did not result in the formation of typical cocoon webs) and long-lasting (spiders from which larvae were removed built similar webs the following night, although some slowly reverted to more normal orbs on subsequent nights). The larva�s ability to induce specific behaviour patterns in the spider indicates that, at some level within the host, such fine behavioural details are independent units, and not artificial constructs. This point is crucial in the use of behavioural patterns as taxonomic characters [8,9]. Many parasites manipulate their host�s behaviour [10�13], but most of them, particularly insect parasitoids, induce only simple changes, such as movement from one habitat to another, eating more or less, or sleeping [12,13]. These changes may be instigated by relatively straightforward mechanisms, for example by the modification of particular receptors [14,15]. Hymenoepimecis�s manipulation of its spider host is probably the most finely directed alteration of behaviour ever attributed to an insect parasitoid. � 2000 Macmillan Magazines Ltd
RELATED ARTICLE:
Figure 1 Webs of the orb-weaving spider
Pleslometa argyra.
William G. Eberhard SimpleToRemember.com - Judaism Online |